


Excursus on Creation of Life and Biological Diversity (Part 32)
October 09, 2019 Time: 30:21Evidence for the Neo-Darwinian Mechanisms
During our last session we examined the evidence for the doctrine of common ancestry, and we saw that the evidence for that thesis is mixed. I think the genetic evidence provides significant support for it, but the fossil evidence seems to tend against it. The absence of transitional forms in the fossil record combined with the evidence of genetics suggests that if the thesis of common ancestry is true then there's something wrong with the explanatory mechanisms of neo-Darwinism. The neo-Darwinian explanatory mechanisms need to be able to give us a good account of both the genetic and the fossil evidence if they are to commend themselves to us.
Before we look more closely at those neo-Darwinian mechanisms, I want to say a word about a couple of questions that were asked last week that I had to beg off on. One of them was if I could think of any transitional forms. You'll remember we distinguished between a transitional form (which is a sort of evolutionary bridge from an ancestral organism to a later organism) and an intermediate form (which is a kind of blend of two different types of organisms). The intermediate forms are not necessarily ancestral to each other. Someone asked: What would be an example of a transitional form? On the way home, I thought of two possible examples. One is an organism called a Tiktaalik. This was a fish that lived about 375 million years ago in the so-called Devonian period. Like the modern lungfish, it was a fish that had the ability to raise itself up out of the water on its bony fins and so spend a brief time in the air. Many evolutionary biologists think that the Tiktaalik is a transitional form between fish and land animals. Another example that is very evident would be the Australopithecines. You can remember this from the expression australo which means “southern” (as in Australia), and pithecus which means “ape.” What these Australopithecines were were bipedal apes that evolved over a million years ago in southern and eastern Africa. Many evolutionary biologists think that humans evolved from these Australopithecines. There are many different kinds of Australopithecines attested in the fossil record, and most of them were probably just dead ends on the tree of primate evolution. They didn't lead to anything further. But many evolutionary biologists think that it may well have been that one of the lineages of these Australopithecines did lead to homo or to man. Those would be a couple of examples of proposed transitional forms in the fossil record. But you’ll remember the point we were making last week was that on the neo-Darwinian theory we're not talking about a few isolated transitional forms; rather there should be millions and millions of these if the neo-Darwinian mechanisms are correct.
The other question that was asked was about the comparison between the genome of chimpanzees and human beings. You'll remember that I remarked on the fact that the genetic sequence of chimpanzees is strikingly similar to that of human beings. They appear to be very closely related. Someone asked me: Are you talking about just that part of the DNA that codes for proteins or are you talking about the entire genome that includes so-called junk DNA as well as this DNA that is coding? The similarity, I can report, between humans and chimps which is genome-wide is 95%. So including the junk DNA, the entire genome, we are about 95% similar to chimpanzees. If you include only the protein-coding sequences (that part of the DNA) the similarity between humans and chimps is 98%. It's striking to compare this with the genetic similarity between mice and rats. Mice and rats seem pretty close together, but in the case of mice and rats, the genome-wide similarity between mice and rats is about 70%. They're about 70% similar in their genome. If you ask just after the coding portion of the DNA then the similarity between mice and rats is about 82%. So 70% and 82% compared to humans and chimps which is 98% and 95%. In other words, the difference between mice and rats genetically is far, far in excess of the difference between human beings and chimpanzees genetically. Now, lest anybody be so foolish as to say, well, then this shows that we're just like apes or that we're just like chimpanzees, what you need to remember is that a difference in the genotype or the genetic structure of an organism can result in monumental changes in the phenotype, that is to say, its appearance and structure and other properties. In other words, all it takes is a change in a few genes and there can be utterly revolutionary consequences in the organisms. So while chimps sit around in forests picking lice off of each other, human beings fly to the moon, write symphonies and novels, build cathedrals, have modern medicine, and ride in nuclear submarines. So don't be deceived by the genetic similarity between chimps and humans into saying we're just like the apes. But that does answer the question that was posed whether or not we're talking about just coding portions of the DNA or the entire genetic sequence.
Before we look at the neo-Darwinian explanatory mechanisms of genetic mutation and natural selection, I think it's worth emphasizing just how extraordinary an extrapolation neo-Darwinism involves. Many of us would probably think that if random mutation and natural selection could explain, for example, the evolution of the horse from a tiny three-toed creature up to a modern thoroughbred today that would really show the power of these neo-Darwinian mechanisms. In fact, evolution within a single kind like that is just nothing compared to the whole range of life. You might think if we could show that random mutation and natural selection could explain, say, how a bat and a whale evolved from a common ancestor that would really show the power of these explanatory mechanisms. Think again!
This figure illustrates the major phyla or divisions of multicellular animals.
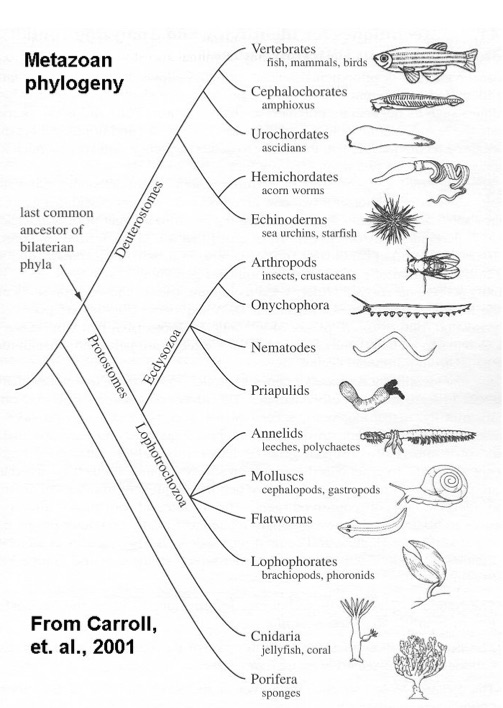
Notice that a bat and a whale are both mammals and vertebrates in the first category – the top category with the picture of the fish. Mammals is just a subcategory of the vertebrates. So even the evolution of a bat and a whale from a common ancestor would be an utter triviality compared to the wide range of the animal kingdom. It would do nothing to explain, for example, how a bat and a sea urchin (which is about the fifth figure down) would be evolved from a common ancestor, not to speak of the difference between a bat and a sponge (down on the bottom of the chart). So the idea that natural selection and genetic mutation are able to explain how everything could have evolved involves an enormous extrapolation – an incredible leap of faith in the efficacy of these neo-Darwinian mechanisms.
If that takes your breath away, get a load at this next chart. This shows you the wide categories of life on Earth.
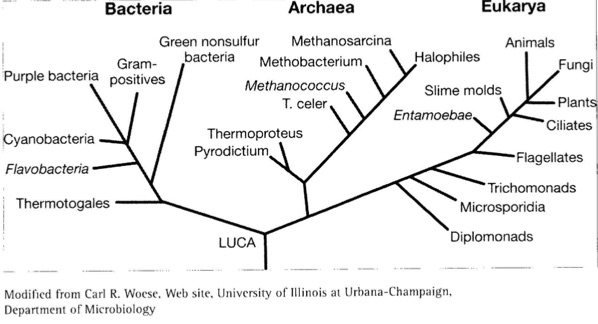
Notice that that entire previous slide that we looked at is contained in that little twig in the upper right-hand corner called “Animals.” I love the modesty of that – “Animals.” That's all of them! Look just to the right of that – “Plants.” The whole plant kingdom is included in that little twig. And these are just eukaryotes – just the twigs on the branch of eukaryotes, which are cells that have membranes containing chromosomes in them. There are still the other two domains containing bacteria and archaea to be accounted for and to come from the LUCA which is the Last Universal Common Ancestor from which allegedly all life arose. The extrapolation of the neo-Darwinian mechanisms from peppered moths and fruit flies and finch beaks to the production and evolution of every living thing on Earth is a breathtaking extrapolation of gargantuan, Brobdingnagian, proportions. And we know that in science such extrapolations sometimes fail. For example, Albert Einstein, having successfully crafted a special or restricted theory of relativity to relativize uniform motion to reference frames, tried to extend or extrapolate the theory of relativity to a general theory of relativity that would relativize all motion including acceleration and rotation. Unfortunately for Einstein, that effort was a failure. The so-called general theory of relativity is really a theory of gravitation. It is the gravitational theory that is Einstein's greatest accomplishment and which replaced Newton's theory of gravity. But Einstein was not able to craft a general principle of relativity that would relativize away accelerated and rotary motion in the way that he had been able to for uniform motion. So that's just one example in science of where an attempted extrapolation in fact failed. Again, I think we're compelled to ask: What is the evidence for the extraordinary extrapolation that is involved in suggesting that these neo-Darwinian mechanisms are responsible for the evolution of all life on Earth from the Last Universal Common Ancestor?
Francisco Ayala, whom I've quoted previously, gives a very typical case of the evidence offered on behalf of these neo-Darwinian mechanisms. Ayala appeals principally to three phenomena. First is the experience of breeders. Secondly is the famous peppered moth experiments. And thirdly would be the development of drug resistance on the basis of random mutation.
With respect to the first of these – the experience of breeders – Ayala appeals to the experience of breeders in producing new varieties of, say, roses or dogs. But such experience obviously does nothing to justify the extrapolation of these mechanisms to the production of grand evolutionary change. Indeed, quite the contrary. The experience of breeders tends to show the limits of these mechanisms. For example, despite decades of effort, breeders have never been able to get chickens to lay more than one egg per day. So breeding actually tends to show the limits of what natural selection and mutation can do rather than their almost infinite plasticity.
Ayala also appeals to the famous peppered moth experiments. But all that happened in that case was that the proportion of light-colored moths in the population decreased and the proportion of dark-colored moths in the population increased. Light-colored moths never evolved into dark-colored moths. It was just a matter of their relative proportions in the population. Taken as evidence of the power of natural selection and random mutation to produce grand evolutionary change, to call such evidence paltry would be to pay it an over-compliment, I think.
START DISCUSSION
Student: I've read a quote many years ago by Dr. Jerry Coyne from the University of Chicago that said that when he found out that the peppered moth experiment was all false, that the moths themselves were glued on the trees, and all of the pictures and everything else, he said it was like being a six-year-old and being told that Santa Claus didn't exist.
Dr. Craig: Coyne is a very aggressive anti-religious evolutionary biologist. That’s an interesting testimony. What you are referring to is that the pictures in the journals were not of the moths naturally lighting on the bark of the dark trees, they were in fact pinned there by the researchers and then the photographs were taken. The moths actually nest up in the leaves.
Student: I emailed him several times. We communicated back and forth for a while until he figured out that . . . I was asking him how did the genes start having dark rather than once I have dark having the proportions change. He's a smart guy. He figured out I was a creationist and stopped.
Student: What weakens the genetic case is that changes in biological organisms are non-random. It's like the cave fish whose eyes have lost their effectiveness. If they're taken out of the cave and put in lighted environments, several generations later they get their eyes back. So the mechanism, the genetic packages, are there and so in the case of the feathered moth it didn't evolve or devolve, it's just that the environment made one variety of the moth more predominant than the other, and then when the environment cleaned up then the mix went back to about 50/50.
Dr. Craig: I'm not familiar with the illustration of the cave salamanders and other creatures that lose sight or color, but if what you're saying is correct it sounds very much like Darwin's finches – that the larger beaks tended to predominate when they were in dry weather (drought) but then during the rainy season the beak proportions would return to what it was before because the information was still there.
Student: Just another note in connection with Denton you mentioned last week. He also made the point that you have in these trees of life you have organisms that have origin in the same branch or what have you but have widely different gestation rates and mutation rates, but yet they're called similar.
Dr. Craig: Yes, I was careful to state my points modestly with respect to the shared genetic code and the similarity of the genetically reconstructed tree of life to a tree of life constructed on the basis of morphology or body plans. It is true that there are places where they don't line up. Turtles, for example, don't seem to fit in well. But those are outliers. The overwhelming pattern, the large-scale pattern, I think is quite supportive of a parallelism, shall we say, between the genetic structure of these animals and their morphological characteristics.
Student: I was watching this nature show which we all do on Sunday evenings a lot of times, and they were talking about the octopus and how different the octopus was compared to any other species. It was amazing transformation the way this octopus could change colors and morph and go into places. But they described it – they said this is more like an alien creature than any other entity on the planet. In other words, it evolved distinctly differently from everything else. And its appearance, the way it changes color and the way that it can morph and go into these tiny places. An invertebrate, in other words.
Dr. Craig: Yes, it's a fantastic animal. The thing that's most noteworthy and famous about the octopus is its eye. The eye of the octopus (in my understanding) is the closest kind of eye to the human eye that exists in nature. The octopus also has a so-called camera eye, and yet obviously it's utterly unrelated – well, it's not closely related shall we say – to human beings on the evolutionary tree. They would be very, very far apart. This is a phenomenon that has been called convergent evolution where it seems like two independent evolutionary pathways both arrived at something very, very similar – in this case, the eye. So that's another puzzle to be explained – how do you have convergent evolution?
END DISCUSSION
I wanted to make the point about the finch beaks that I just mentioned in regard to an earlier question. On the Galapagos Islands when Darwin visited those, he measured finch beaks of different sizes of the birds that were there. But, again, like the peppered moths, nothing really evolved here. Again, it was just that the proportion of the finches with large beaks increased during the dry season (during the times of drought) and then the proportions with small beaks would decrease during those droughts. But when the rains would come then the normal proportions of the beaks would return among the population of finches there. So it wasn't as though the finches were evolving into something else. It was just a matter of decreasing proportions in the population.
Ayala also mentions in this connection the speciation that occurs among fruit flies in the Hawaiian Islands. The Hawaiian Islands are extremely isolated and so tend to be sealed off from outside influences. Yet, 500 species of fruit fly exist in the Hawaiian Islands. This is 1/4 of all the species of fruit fly that exists in the entire world. It would seem bizarre to think that God somehow favored fruit flies in the Hawaiian Islands that he would create 1/4 of all the world's species of fruit flies in Hawaii. The evidence, I think, would more persuasively point to their common ancestry and evolution. I think we could agree that this is well within the limits of what neo-Darwinian mechanisms can achieve. It could produce a multiplicity of different fruit flies on the various islands. This is probably due to genetic drift of the flies among the islands – a non-adaptational change in them in these various populations of flies on the islands. But, again, that hardly would go to justify the extrapolation of these mechanisms to the grand evolutionary scenario. It's a pretty big leap from fruit flies to explaining the entire history of life on this planet.
Finally, Professor Ayala appeals to the ability of organisms to develop resistance to drugs and poisons through random mutation and natural selection. He points out how an unacceptably improbable double mutation can occur one step at a time to produce cumulative change. Then he extrapolates this process to explain vast evolutionary change. But of course the question is precisely: Can the example of adaptation or resistance to drugs and poisons be extrapolated in that way?
In his book, The Edge of Evolution, Michael Behe argues that, in fact, such an extrapolation is illegitimate. He maintains that the very evidence of organisms’ development of drug resistance is a powerful indication of the limits of random mutation and natural selection to evolutionary change.
He uses the example of malaria and the human immune system. Malaria and the human immune system have been waging war against each other for over 10,000 years. Since the advent of modern medicine human beings have been developing anti-malarial drugs to try to destroy this organism. Unfortunately for us, the malarial population is huge. The average person infected with malaria has over one trillion malarial cells in his body. Therefore malaria mutates extremely rapidly and so has been able to develop resistance to every drug that we've hurled at it. Simple single point mutations are enough to make malaria drug-resistant. For example, one mutation in an amino acid at point 108 suffices to render malaria drug-resistant to the drug pyrimethamine.
On the other hand, Behe points out there is tremendous selective pressure for the human immune system to develop some sort of a defense against malaria, but it hasn't been able to do so. Instead, what's happened is that a mutation has occurred in the human respiratory system – not in the immune system, in the respiratory system – which makes some people immune to malaria, namely sickle hemoglobin. Unfortunately, the downside is that it also produces sickle cell anemia which is eventually deadly.
This is where things get really interesting. Despite its incredible mutation rate which has enabled malaria to overcome every drug we've thrown at it, malaria has never in all those thousands of years and trillions of mutations been able to overcome sickle hemoglobin. Molecular biology explains why. Resistance to a drug can result from a simple single-point mutation. But overcoming sickle hemoglobin would require multiple simultaneous mutations or else a sequence of mutations occurring blindly, and both are just too improbable to occur. So we see the limits of random mutation and natural selection in malaria's inability to overcome sickle hemoglobin in the human respiratory system.
We're out of time, so let's close there. Next time we'll look at Behe’s example of how HIV provides another example – another case study – in addition to malaria.[1]
[1] [1]Total Running Time: 30:21 (Copyright © 2019 William Lane Craig)